
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|

|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||

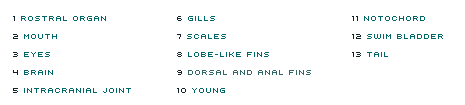
In 1836, the eminent naturalist Louis Agassiz first described a fossil fish he called Coelacanthus (Greek for "hollow spine"). Since then, fossils of some 125 species of coelacanth have turned up, dating from about 400 to 66 million years ago. After that, they vanish from the fossil record and so were thought to have gone extinct 66 million years ago. Then, in 1938, a living coelacanth was caught off South Africa (see Moment of Discovery). Since that historic find, many other individual coelacanths, including specimens of a new species in Indonesia, have come to the surface, enabling scientists to carefully study this most unique creature. Using our clickable coelacanth, see what makes this fish stand out from all others. Note that this is only a cursory treatment; for more on coelacanth anatomy and physiology as well as ecology and behavior, see the books listed in Links & Books.—Peter Tyson 
This large, jelly-filled cavity in the center of the snout is thought to be an electrosensory device for detecting weak electrical impulses given off by prey. Evidence for this function first came from studies of the organ's anatomy and its nerves as well as the structure of the brainstem. Later, experiments conducted from a submersible confirmed that coelacanths can detect and respond to electrical fields in the water, strongly implicating the rostral organ for this role. Like the intracranial joint, this feature is unknown in any other extant animal.

With the aid of the intracranial joint and other cranial muscles, the coelacanth usually swallows its prey whole. Its teeth are designed not so much to grab or slice fish but to prevent them from escaping once the coelacanth has sucked them into its gaping maw. Beneath the chin lie large, bony plates.

The coelacanth has large eyes attached to thick optic nerves. Each eye has few cones, which register color, but many rods, which detect light. So the coelacanth is virtually color-blind but can see extremely well in the dim light of the great depths where it lives. This ability to see in near darkness is enhanced by a layer behind the retina called the tapetum lucidum. Similar to that found in domestic cats, the tapetum acts like a mirror, intensifying weak light.

The small brain occupies only about 1.5 percent of the braincase in mature individuals. Its weight is negligible: in a 90-pound coelacanth, the brain weighs less than a tenth of an ounce. (Adult human brains typically weigh about three pounds.) No known living vertebrate has so miniscule a brain in relation to its body size. Yet such a rudimentary brain has clearly done nothing to hinder the species' long-term survival.

Previously known only from fossils of primitive fishes, this joint allows the coelacanth to open its mouth especially wide in order to swallow fish and other prey. A pair of powerful muscles—the largest muscles in the head—cross the intracranial joint and are thought to allow the coelacanth to powerfully grip prey fishes between its teeth.

The coelacanth has gills similar to those of fish living at the depths in which it is commonly found (500 to 800 feet). The gills have a relatively small surface area compared with the body mass of the fish; these are the gills of a slow-going rather than an active fish.

The scales are hard, woven tight as armor, and rough to the touch. The roughness comes from tiny, tooth-like spikes called denticles, which provide the coelacanth with protection against rocks and predators. The scales are a steely blue-gray in color and bear irregular white spots, the patterns of which are distinctive enough to enable researchers to identify individuals.

Unlike most living fishes, the two living species of coelacanths, along with six living species of lungfishes, belong to a group known as the lobe-finned fishes. Projecting from the body on stalks rather than attaching directly to the body, these fishes' paired lobe fins are supported by the same basic bones as your arms and legs. Coelacanths even move their paired fins much like land animals move their limbs: the right pectoral fin moves in conjunction with the left pelvic fin, for example. And their movement is extremely dexterous. They scull the water like oars and can rotate through 180°.

In addition to their paired fins, the second dorsal and anal fins also have stout, muscular lobed bases. The first dorsal fin, however, lacks such a lobe, but it can be raised or lowered like a fan to change its area.

The coelacanth is ovoviviparous, meaning that eggs are fertilized internally and are retained in the mother's oviduct so that the young are born alive. Pups—26 were found in one mother—develop from huge eggs that are among the largest vertebrate eggs known. The growing young live off a large yolk sac until birth. At this time they are about a foot long and look like miniature versions of their parents.

Elastic and fibrous, this thick-walled tube filled with oil serves as a kind of backbone. In most creatures with backbones, vertebrae replace the notochord in the embryonic stage of development. But the coelacanth has no fully formed vertebrae, and so the notochord provides its longitudinal support.

This lies beneath the notochord, embedded in fat. In most fish, the swim bladder is filled with air, but in the coelacanth, it is packed with oil and fat. Both types of bladder have a similar purpose, however—to increase buoyancy.

The tail is flat and powerful, enabling the coelacanth to bolt
forward in great thrusts to attack prey or escape predators. It is
divided into three sections, with a small tail fin in the middle.
(Coelacanth discoverer Marjorie Courtenay-Latimer called this
secondary tail fin a "puppy dog tail.") Able to both rotate and flex
from side to side, this so-called epicaudal fin is thought to help
the coelacanth with trim and balance.
|
||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||||||||||||||||||||||||||||||||||||||
|